
小林 芳郎
マクロファージ-死細胞を取り込んだマクロファージの応答-
私たちの研究 (2) -未熟樹状細胞と死細胞-
さて成熟樹状細胞(下図)は抗原提示にすぐれ、免疫応答に重要な働きをしていることで知られる。
一方未熟樹状細胞はマクロファージほどではないがアポトーシス細胞を貪食する。
そこでMTは骨髄由来未熟樹状細胞を用いて、後期アポトーシス細胞(ひとによっては2次的ネクローシスともよぶ)と共培養したときに初めて産生されるサイトカインを探索した。するとIL-6とIL-12p40がmRNAレベルでもタンパク質レベルでも産生された。これらはネクローシス細胞(1回凍結融解を起こさせて得た細胞)との共培養では産生されなかった。産生量の多いIL-6についてさらに詳しく検討した結果、これが未熟樹状細胞を未熟な状態にとどめる作用をもつことがわかった(→Cell Immunol 226, 105-115, 2003, 228, 138, 2004)。この機構と、アポトーシス細胞を貪食した未熟樹状細胞が未知の機構によりLPS刺激による成熟化を起こしにくくなることとによって、アポトーシス細胞が生体に現れたときに免疫応答が伴わないようにしているのではないかと考えられる。
未熟樹状細胞はしばしばマクロファージと近接して観察される(J Exp Med 183, 1865-1878, 1996; Int Rev Cytol 197, 83-136, 2000)。
そこでMTはチオグリコレート誘導腹腔マクロファージと後期アポトーシス細胞(ひとによっては2次的ネクローシスともよぶ)とを共培養するときに未熟樹状細胞を共存させてMIP-2産生に対する効果を調べた。すると未熟樹状細胞の共存によってMIP-2産生が抑制され、トランスウェルで細胞の接着を遮る([DC+Apo]の[ ]がそれをさす)とその効果が失われた(下図、赤枠)。つまり未熟樹状細胞は接着依存的にマクロファージのMIP-2産生を抑制しているようである。さらにKupffer 細胞でも、さまざまな後期アポトーシス細胞でも、同様の結果を得ているので、この現象は一般的だと考えられる。
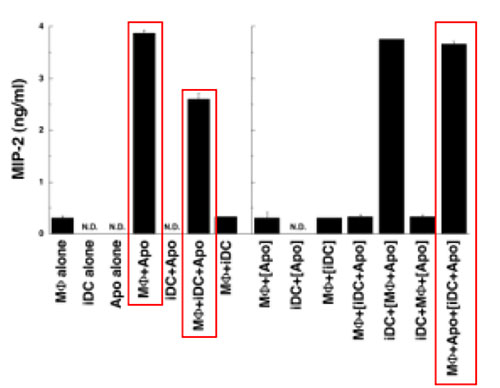
抑制の機構として容易に想像できるのは、アポトーシス細胞の奪い合いだが、検討したところ、マクロファージの貪食はむしろ促進されていた。そこで抑制性サイトカイン(TGF-βとIL-10)の抗体を用いたところ、抑制が解除された。これらのサイトカインがどの細胞から産生されているのかをmRNAレベルで調べた(下図)。なおこれらのサイトカインはタンパク質レベルでは検出限界以下だった。
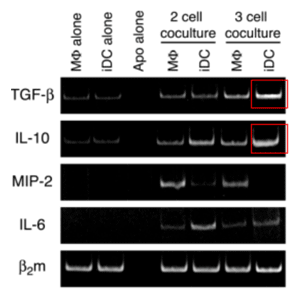
結果、3者共培養によって、未熟樹状細胞からのTGF-βとIL-10産生が促進された。これにより、マクロファージからのMIP-2産生が抑制されたと考えられる。以上の結果は、アポトーシス細胞の取り込みに伴う応答の新たな制御機構を明らかにしたものといえる(→J Leukoc Biol 75, 865-873, 2004)。MTは2004年3月これらの業績により理学博士の学位を取得した。
![]()