カメとカエルに学ぶ野外生態学
 生息地の破壊や温暖化などの地球規模での環境変動、あらたな病原微生物の出現、そしてさまざまな化学物質の悪影響により、世界各地で両生類の減少が進行している。日本においては、都市化による水田の減少、圃場整備による乾田化といった生息地の減少が
両生類個体群の主な減少要因となっている。
生息地の破壊や温暖化などの地球規模での環境変動、あらたな病原微生物の出現、そしてさまざまな化学物質の悪影響により、世界各地で両生類の減少が進行している。日本においては、都市化による水田の減少、圃場整備による乾田化といった生息地の減少が
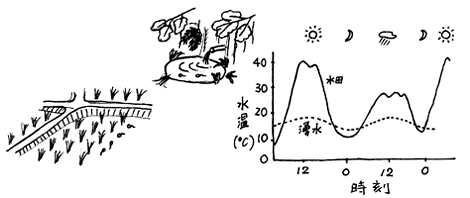
両生類個体群の主な減少要因となっている。 カエル類は、皮膚からの水分蒸発にともなう気化熱を利用して、体温の上昇を防いでいる。蒸発によって失われる水分は下腹部の皮膚を通して基質(湿った地面など)から吸収することで、補っている。そのため、高温かつ乾燥した条件下にあるカエルにとって基質からの水分補給は不可欠である。このようなカエルの生理的特長を考慮するならば、温度と水分条件の面から復元された生息環境の適切さを評価することが可能ではないだろうか。
カエル類は、皮膚からの水分蒸発にともなう気化熱を利用して、体温の上昇を防いでいる。蒸発によって失われる水分は下腹部の皮膚を通して基質(湿った地面など)から吸収することで、補っている。そのため、高温かつ乾燥した条件下にあるカエルにとって基質からの水分補給は不可欠である。このようなカエルの生理的特長を考慮するならば、温度と水分条件の面から復元された生息環境の適切さを評価することが可能ではないだろうか。地理生態学研究室における評価計画
1.変態したカエル類の生息地評価
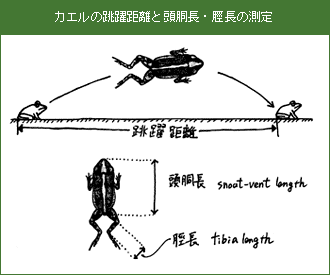
- 身近なカエルの野外における活動体温を知る
- 活動体温とカエルをとりまく外界温度との関係を明らかにする
- カエルの個体レベルにおける生理特性と体温との関係を明らかにし、生理的に最適な体温の範囲を知る
- カエルの体温と体表面からの水分蒸発の関係が、変動する外界環境の諸パラメータのもとでどのように変化するのかを明らかにする
- カエルの個体レベルにおける生理特性と水分含有量との関係を明らかにし、カエルが耐えうる脱水状態の限界を知る
- センサーに用いるカエルモデルの素材を検討する
- 開発したセンサーを用いて、カエルの減っている場所とそうでない場所で生息地の評価を行う
- カエルの再導入を企画している環境をこのセンサーで評価する
- 生息地として適していると判定されたビオトープに卵塊を再導入して、翌年繁殖に戻ってきた個体の数によって、実際の評価を行い、センサーによる評価を裏付ける
- センサーの実用化と普及啓蒙に向けた企画を策定する
2.水中で生活する幼生の生息環境評価
- さまざまな温度環境におけるオタマジャクシの環境選好性を温度勾配装置を用いて明らかにする
- 生理的な致死的限界温度を知る
- 選好温度範囲における生理的活性(運藤能力、摂食性など)の温度依存性を明らかにする
- 多数の温度データロガーを用いて、温度環境の空間変異を明らかにする
- オタマジャクシの種類別に、温度から見た生息適地の広がりを空間的に明らかにする
平成13年度に発足したこのプロジェクトは、まず卒業研究として初期の成果、上記計画のうち1−1,2,6をあげることができた。14年度は、引き続き卒業研究として、カエルの生理的特性と体温の関係を明らかにした上で、カエルモデルのセンサーを野外に設置し、具体的な環境評価を試みた。15年度は。1−5に関する実験を行って、カエルモデルの水分蒸発特性とカエルそのものの水分蒸発特性を比較検討した。16年度は、千葉県に生息するカエル類の幼生を材料に、種別の温度選好性を実験的に明らかにしたうえで、野外における繁殖場所評価につなげていく。近い将来には、学校ビオトープ等を利用して具体的な環境評価を行い、生息環境の復元に向けて努力していく。
実習の目的
本実習は、カエルの幼生(オタマジャクシ)と変態個体を対象として、野外における生息環境の物理的諸条件(流速、温度など)を把握して、彼らの生息場所選択を形態的特徴、運動能力、温度選好性と関連させて理解することを目標とする。
1.セミナー
「カエル類の個体群存続可能性分析と生息地適合性評価手法の開発
−変態個体の体温調節行動に注目して−」 TA 中村 有(修士2年)
アカガエル類を対象とした保全生態学的研究について、修正課程の研究成果と計画を実習内容と関連させた話題提供。
2.実習の構成
- 森林、渓流、湧水、水田、池、湿地などにおけるカエル類とその幼生の生息環境の把握
- 自記記録温度計を用いた野外におけるカエル幼生の温度環境測定
- カエル(スポンジ)モデルとヘビ(パイプ)モデルを用いた作用温度の測定
- 温度勾配装置を用いたカエル幼生の温度選好性実験(2種類以上の比較)
- 河川渓流におけるカエル幼生の遊泳能力測定
- データシートの作成、調査、データ整理、解析、レポート作成
1.カエル類とその幼生の生息環境の把握
1.カエルの捕獲、体温測定、幼生の生息環境の把握
 調査地(市原市石塚、浦白川源流域)において、カエル類の生息状況を調査する。
調査地(市原市石塚、浦白川源流域)において、カエル類の生息状況を調査する。
カエルを発見したら、最初にカエルがいた場所の環境を記録する。次に、その場でカエルを刺激し(枝などで)跳躍させる。着地した個体をすばやくクモ網で捕獲し、サーミスタ温度計を総排泄孔に挿入し、体温を測定する。その直後に、基質(カエルがいた場所の地面、石の上など)の温度、日陰の気温、最寄の水場の水温を測定する。その後、カエルの頭胴長と脛長をノギス(0.1mm単位)で、体重を簡易電子天秤(0.1g単位)で測定し、捕獲地点に放逐する。変態したばかりの子ガエルは脆弱で、高温に弱いので、この測定には用いない。
2.カエル幼生の生息場所
調査地に生息するカエルのうち、実習時期(6月中旬)に幼生が観察できるのは、カジカガエル、モリアオガエル、シュレーゲルアオカエル、ニホンアマガエル、ツチガエルの5種である。この5種の幼生がどのような環境に生息しているのか、実習地をくまなく探して、種ごとの生息場所を突き止める。
幼生の種類と発生段階を記録し、生息環境を分類(水田、用水路、防火用水、プール、湧き水、池、ため池、渓流)記録する。水深、底質の種類(泥、砂、落ち葉、砂利、岩)、水温(表面と底)、気温。溶存酸素、電気伝導度、pHを測定記録する。流れのある場所では、流速を浮き流し法(浮きを一定の距離流して、時間をストップウォッチで測り、流速を求める)によって測定する。
3.カエル幼生の形態的特長
 各種のオタマジャクシを最低核0個体捕獲し、5%ホルマリン溶液で固定標本を作成する。宿舎に持ち帰り、形態測定を行う。測定箇所は、幼生の形態的特長を比較し、種の形態的特長をうまく表現できるような形質を選び出すこと。
各種のオタマジャクシを最低核0個体捕獲し、5%ホルマリン溶液で固定標本を作成する。宿舎に持ち帰り、形態測定を行う。測定箇所は、幼生の形態的特長を比較し、種の形態的特長をうまく表現できるような形質を選び出すこと。
2.自記記録温度計を用いた野外におけるカエル幼生の温度環境測定
 各種の幼生が発見された水環境を種類ごとに3ヶ所選び(系5測定点)、水温の継続測定をデータロガー(自記温度記録計)を用いて行う。水温の測定期間は、6月11日の午後から14日の午後(3.5日)とし、記録インターバルは20分間隔とする。得られる総データ数は1地点薬230(3x24x3.5)となる。さらに、カエル幼生が全く発見できなかった水環境を3ヶ所選び、そこにもデータロガーを設置し、幼生が利用していない水界の温度環境測定を行う。
各種の幼生が発見された水環境を種類ごとに3ヶ所選び(系5測定点)、水温の継続測定をデータロガー(自記温度記録計)を用いて行う。水温の測定期間は、6月11日の午後から14日の午後(3.5日)とし、記録インターバルは20分間隔とする。得られる総データ数は1地点薬230(3x24x3.5)となる。さらに、カエル幼生が全く発見できなかった水環境を3ヶ所選び、そこにもデータロガーを設置し、幼生が利用していない水界の温度環境測定を行う。
3.カエル(スポンジ)モデルとヘビ(パイプ)モデルを用いた作用温度の測定
変態したカエルとその捕食者である蛇の生息環境を評価するために、スポンジモデルとパイプモデルを用いた作用温度測定を行う。作用温度とは、動物の体温を物体の熱収支によって近似するもので、カエルを蛇の体温を模倣する素材として選ばれた水分70%の切り花用スポンジと銅製のパイプ(直径が蛇の胴直径に等しく、長さがとぐろを巻いたヘビのトグロの直径に等しくなるように切り揃え、灰色に塗ったもの)を用いる。
予め作成しておいたスポンジモデル5個とパイプモデル5個にデータロガーを装着し、日当たりのよい乾燥した草地から、森林内にかけて並列して設置する。この装置によって、日向と日陰におけるカエルとヘビの模擬体温測定を行う。温度の測定間隔は、10分毎とし、11日の午前中から14日の午後まで約3.5日設置、測定を行う。
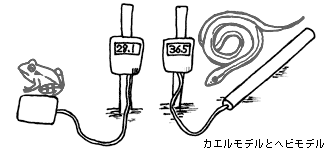 モデルの設置場所は、以下の5通りとする。
モデルの設置場所は、以下の5通りとする。
- 日当たりのよい乾いた草地(田の土手)
- 日当たりのよい湿った草地(休耕田の湿地)
- 林縁
- 湿った林床
- 渓谷沿いの河原
4.温度勾配装置を用いたカエル幼生の温度選好性実験(2種類以上の比較)
1.装置の準備
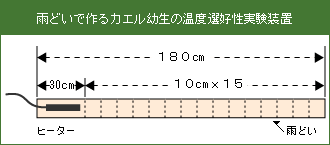 180cmの雨どいの端から30cmを測り、残りの150cm部分を10cmごとの区画に分けて目印をつけ、区画の中央にも目印をつける。雨どい全体い1リットル程度の砂利を均一に敷き詰め、1.5cm程度の水深になるように水を張る。10cm区画を作らなかった30cmの部分にヒーターを千知するが、ヒーターは設定温度が調節されているので上部の調節つまみは動かさない。また電源を繋いだ状態でヒーター全体が水中に浸かるように、おもりをヒーターの両端に置いてからヒーターを作動させる。温度勾配が出来上がるまで1時間程度かかる。
180cmの雨どいの端から30cmを測り、残りの150cm部分を10cmごとの区画に分けて目印をつけ、区画の中央にも目印をつける。雨どい全体い1リットル程度の砂利を均一に敷き詰め、1.5cm程度の水深になるように水を張る。10cm区画を作らなかった30cmの部分にヒーターを千知するが、ヒーターは設定温度が調節されているので上部の調節つまみは動かさない。また電源を繋いだ状態でヒーター全体が水中に浸かるように、おもりをヒーターの両端に置いてからヒーターを作動させる。温度勾配が出来上がるまで1時間程度かかる。
2.温度選択性の記録
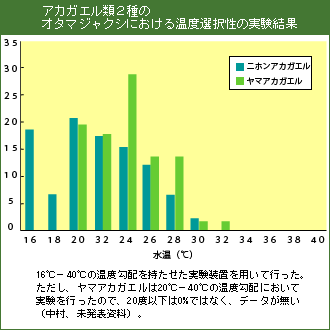 最初に、各区画に温度勾配ができているか温度計を用いて確かめる。区画の温度を測定する時は、区画の中心点の温度を測定するものとし、推進は中層と底層の中間とする。温度勾配が確認できたら、1回の実験につきオタマジャクシ10匹を、各区画のうち中央の区画に放し、その後のオタマジャクシの行動を観察し、1分後、3分後、10分後にオタマジャクシの存在した区画と各区画の温度を記録する。オタマジャクシを回収した後、次の10匹について同様に実験・記録を行う。アマガエル・シュレーゲル、モリアオガエル各20匹について実験を行う。なお、ヒーターを作動させた後、4−5時間以上経過すると温度勾配が顕著に崩れてくるので留意する。
最初に、各区画に温度勾配ができているか温度計を用いて確かめる。区画の温度を測定する時は、区画の中心点の温度を測定するものとし、推進は中層と底層の中間とする。温度勾配が確認できたら、1回の実験につきオタマジャクシ10匹を、各区画のうち中央の区画に放し、その後のオタマジャクシの行動を観察し、1分後、3分後、10分後にオタマジャクシの存在した区画と各区画の温度を記録する。オタマジャクシを回収した後、次の10匹について同様に実験・記録を行う。アマガエル・シュレーゲル、モリアオガエル各20匹について実験を行う。なお、ヒーターを作動させた後、4−5時間以上経過すると温度勾配が顕著に崩れてくるので留意する。
実験に用いる幼生の発生段階はGosner(1960)のステージ(35−39)にそろえる。
温度勾配実験装置について
私たち地理生態学研究室では、カエルの保全と目標とした研究を進める上で、カエルと温度環境との関係を重要なテーマの一つとして考えている。そのため、様々な種の温度選択性を検討することが必要だが、野外において性格にその選択性を調査、同定することは難しい。そこで研究室内で簡便かつ正確に温度選択性を調査するために、私達は温度勾配実験装置の開発、改良を行ってきた。
これまでは本実験のようにヒーターによる過熱だけで温度勾配をつけていたが、さらに冷却装置を反対側に設置して現在も実験を継続中である。現在はオタマジャクシの温度選択性を検討しているが、さらに他の水生生物でも同様の実験が行えるだろう。
5.河川渓流におけるカエル幼生の遊泳能力測定
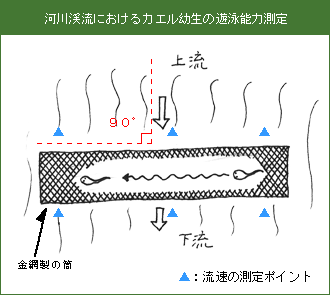 今回、実習地で観察できるカエル幼生5種のうち、カジカガエルが流水性で、モリアオガエル、シュレーゲルアオガエル、ニホンアマガエルの3種は止水性、ツチガエルはどちらの環境にも見られる。流水性のカエル幼生は、止水性のカエル幼生よりも流れに抗して遊泳する能力に優れていると考えられるので、いろいろな流速の下での遊泳速度を測定し、5種の遊泳能力を評価する。
今回、実習地で観察できるカエル幼生5種のうち、カジカガエルが流水性で、モリアオガエル、シュレーゲルアオガエル、ニホンアマガエルの3種は止水性、ツチガエルはどちらの環境にも見られる。流水性のカエル幼生は、止水性のカエル幼生よりも流れに抗して遊泳する能力に優れていると考えられるので、いろいろな流速の下での遊泳速度を測定し、5種の遊泳能力を評価する。
遊泳能力は、金網製の筒(長さ1m、直径10cm)を流れに直角に置き、その中に入れたカエル幼生を流れに抗して泳がせ、その遊泳速度を測定する。測定地点の流れの速さは、流速計を用いて筒の上流側と下流側について、両端と中央部で測定する。カエル幼生が体験する流速は、この6測点における流速の平均値によって代表させる。
測定は、流れの急なところ、緩い所、ない所、3地点で行い、1地点で1種につき、10個体の遊泳速度を測定する。
6.データシートの作成、調査、データ整理、解析、レポート作成
第2部 淡水性カメ類の生態学
はじめに
 カメは万年、ツルは千年、という言葉があるように、カメは長寿の代表的生物である。分類学的には、脊椎動物門、爬虫類網、カメ目に属する生物の総称で、約2億年前の三畳紀末に出現し、白亜紀末の大規模な絶滅をもくぐり抜け、現在では地球上に約230種が知られている。同じ爬虫類に属するトカゲ類の約4000種、ヘビ類の3000種に比べると種数では見劣りするが、装甲をまとったユニークな体、海洋環境から砂漠までさまざまな環境に進出した適応性は、生物進化の観点から大変興味深い生物であると言える。
カメは万年、ツルは千年、という言葉があるように、カメは長寿の代表的生物である。分類学的には、脊椎動物門、爬虫類網、カメ目に属する生物の総称で、約2億年前の三畳紀末に出現し、白亜紀末の大規模な絶滅をもくぐり抜け、現在では地球上に約230種が知られている。同じ爬虫類に属するトカゲ類の約4000種、ヘビ類の3000種に比べると種数では見劣りするが、装甲をまとったユニークな体、海洋環境から砂漠までさまざまな環境に進出した適応性は、生物進化の観点から大変興味深い生物であると言える。
身近な湿地環境にたくさん催促するカエル類と同様に、子供の頃にカメと接したことのない人は少ないのではないだろうか?けれども、縁日の夜店で売られていたゼニガメが子供たちの人気の的であった割には野生のカメを見た経験がある人は意外と少ない。では、野生のカメは人目に付かないほど少ない生き物なのだろうか?
答えは否である。実は千葉県の河川には驚くほどたくさんの淡水性カメ、イシガメとクサガメが生息している。しかし、このことは意外に知られていない。そのため、開発行為に先立って行われる環境アセスメントなどにおいても、淡水性カメ類に木を配った丁寧な調査が行われたことはほとんどなく、彼らの存在は十分に知られることなく、住み場所を奪われてきた。
そこで、今回は淡水性カメ類の生態と保全とテーマとして、実習を実施する。実習地として選んだ君津市福野、怒田は、房総丘陵の大福山山系のほぼ中心部に位置し、房総丘陵全体をみてもその中央部に位置している。大福山は、東京湾に注ぐ養老側の3つの支流(浦白川、芋原川、梅ヶ瀬川)と小櫃川の1つの支流(御腹川)の水源地となっており、福野小学校の北東に位置する浦白川源流部と南東に位置する梅ヶ瀬渓谷は、千葉県の自然環境保全地域の指定を受け、将来的な開発から守られている。その一方で、小櫃川水系に属する御腹川の源流部には大小4つの産業廃棄物最終処分場が既設、あるいは建設中であり、千葉県の自然環境の良い面と悪い面を併せ持った立地環境となっている。
これらの流域は、水田耕作が継続している里地・里山的環境(御腹川流域、芋原川流域、浦白川流域)としでに耕作が放棄され原生的自然への復帰が図られている流域(梅ヶ瀬川流域)に分けられ、房総半島の原生的自然と里山としての2つの側面をコンパクトに体感できる立地となっていることが大きな特徴である。
長寿のカメは、実験室での生理学的研究には向いていないが、地域に根ざして長期にわたって自然環境を監視していく上での素材として、数々の利点を備えている。今回の野外実習を通じて、淡水カメ類の生態調査法や保全生態学を学んでほしい。
1.千葉県の淡水性カメ類の生息状況
千葉県では、今までに3種の在来種(イシガメ、クサガメ、スッポン)と複数の外来種(ミシシッピアカミミガメ、キバラガメ、ミナミイシガメ、ヨツメガメ、カミツキガメ、ワニガメ)などが記録されている。これらの種類のうち、在来種の分布情報は千葉県立中央博物館で収集・整理が進んでいる。外来種のうち、カミツキガメに関しては印旛沼水系を中心に調査が進行中である。
各河川水系におけるカメ類の詳細な分布調査は、これまでに館山市佐野川、富津市亀沢、君津市小糸川、市原市村田川、千葉市都川、千葉市花見川、佐倉市鹿島川、白井市二重川などの諸河川で実施されており、全国的にみても千葉県は淡水性カメ類の野外調査が密度濃く行われている地域であるといえる。
これらの調査によって、在来種と外来種の分布状況は以下のように整理されつつある。すわなち、イシガメは南部の丘陵地の河川中上流部を中心に、クサガメは北総地域の平野を流れる河川を中心に分布しており、外来種であるミシシッピアカミミガメやカミツキガメは都市化の進んだ河川を中心に分布している傾向がある。
2.淡水性カメ類調査法
カメは、捕獲が容易で、固い甲羅を測定するため、測定値の個人差が出にくく、しかも容易に年齢を知ることができるので、個体群の諸パラメータを得ることがそれほど難しくない動物である。そこで、動物個体群の調査方法を学ぶ材料として非常に適しており、本実習ではカメを対象とした野外調査を行う。
1.淡水性カメ類の生活史研究概観
野外における淡水性カメ類の生活史研究は、主にアメリカ合衆国のアメリカヌマガメ、ドロガメ、カミツキガメ類を対象とする研究によって進展した。特に、ミシガン大学の管理する湿地、ジョージア州立大学の附属研究所が所轄するサバナ川流域の生態系保護区での研究が著名である。しかし、カメ類が長期の個体群研究に適した材料であるため、これら以外の大学でも研究が盛んに行われてきた。
カメ類の研究が生物学全般に与えたトピックの1つに、温度依存性決定の発見がある。性染色体を持たないカメ類の性は、卵の中の胚が経験する温度、すなわち卵が産み込まれた土中の温度に左右されるが、性染色体を有するグループでも温度依存性決定が行われていることがわかった。性染色体は、雄ヘテロ、雌ヘテロの両方の場合が存在する。
- 野生のカメ個体群の人口統計学的解析で必要とされる変数
- 生息個体数
- 年齢構成
- 一腹卵数(1個体のメスが1回に産む卵数)
- 一シーズンの産卵回数
- 性成熟・繁殖開始年齢
- 各発育段階毎の年間生残率
2.カメの野外調査方法
1.カメはワナで捕獲する
カメは水生生物で、生活の大半を水中ですごすため、にごった水中にいる姿を確認するのは困難である。さらに、夜間に活動する傾向が強いので、カメを直接目でみつけて捕まえるのは難しい。
カメがいる川かどうかは、春先に川の土手を歩いて確かめる。水温が低いこの時期には、カメたちは川岸でよく日光浴をする。1匹でも日光浴を行っている個体が見つかれば、その川にはかなりのカメがいると判断できる。しかし、日光浴をしているカメを捕まえるのは至難の業である。素早く、川岸をすべり下りて、水中に姿を隠してしまうからだ。一度水中に逃げたカメを追いかけるのはとても難しく、また危険である。
カメは夜になると川から出て、餌を探すために水田や湿地を歩き回る。すると、田んぼの泥の上にはカメの足跡が残される。足跡の間隔は甲羅の幅よりも少し広めで、中心には尻尾が泥をなぞったスジが残される。カメの足跡が残された田んぼを見つけたら、夜間に田んぼの中を懐中電灯で照らしてみよう。きっと、カメがみつかるだろう。とはいえ、稲の植えられた田にむやみに立ち入ってはならない。
日光浴をしているカメや、足跡をみつけたら、その近くの川でワナかけをして捕獲調査を行う。ワナは効率がよくカメを捕獲する道具で、篭(カゴ)ワナという形式のものを使用する。針金や鋼鉄製の外枠に漁網をはり、入り口は漏斗状になっていて一度入ると出られない構造になっている。カメをワナの中におびき寄せるために、魚の頭や内臓を誘引餌として、ワナの中に入れる。
餌を入れたワナを川に設置する。ワナの入り口は下流に向ける。餌のエキスが下流に流れ、それにそって亀が誘引されることが期待される。しかし、誘引効果を高めようとして流れの速い場所に設置すると意外にカメがかからない。川の流れが速すぎると捕獲効率が落ちてしまうようだ。ワナの一部が水面に出るように位置を調節しなければならない。ワナ全体が水没してしまうと、カメは空気呼吸ができずに窒息死してしまう。また、雨で増水した場合にはたとえ設置時にはワナの一角が水面上にあったとしても、増水で水没してしまう危険が高い。天気予報に気を付けて、増水の危険があるような日には、無理に調査を決行してはならない。増水によってワナがカメごと流されてしまう危険性もあるので、ワナにはロープを数メートル分くくりつけておき、そのロープを陸上の木の幹や枝、人工の柱にくくりつけておく。
ワナは夕方に設置し、翌日の午前中に引き上げてカメが捕獲されているかを確認する。ワナをかけた場所を地図に記入し、設置時間を書き留める。その後、ワナをしかけた場所の環境を記述する。設置位置は、地図に記録し、緯度経度をGPSで読み取る。一般的な水質測定(水温をかならず記録し、pHなどは随時)に加えて、川の形、川幅、水深、岸の高さ、水温、気温、天候、流速など、カメの生活に影響を与えそうな要素について記録を取る。
2.捕獲したカメの取り扱い
初めてワナを川にしかけた番は、期待でなかなか寝付けない。しかし、寝不足の状態で川に行くと、足を踏み外して転落したりして危険なので、しっかりと睡眠をとる。きっちりと目覚めたら、期待に胸を膨らませて、ワナをかけた場所に行く。岸からワナをのぞき込んでもカメが獲れているかどうか確認が難しいが、水面に出ている部分にカメが体を見せている場合もある。そういう場合は大漁間違いない。
岸の竿や樹木にくくりつけたロープをたぐり、ワナを引き上げる。時刻を記録する。カメが入っていてもいなくても、一度岸にワナを引き上げる。開けた見通しの聞く場所で、ワナをはずし、中のカメを取り出す。複数のワナを仕掛けた場合には、ワナに番号をつけておく。取り出したカメを入れる袋は、洗濯用のネットを使う。少し使い古したものがネットの網目が柔らかくなっているので、カメの皮膚を傷つけないのでよい。ネットのチャックを広げ、カメを一匹一匹袋の中に入れる。捕獲されたカメの種類と個体数を記録用紙に記入する。カメの測定は実験室かベースキャンプに持ち帰って行うほうが良い。
ワナを何ヶ所にもかけた場合には、それぞれの場所でとれたカメをカメの入った袋ごと衣装ケースなど大きめの容器に入れる。油断すると、袋に入ったままの状態ではカメは移動して視野から消えてしまう。もし、その状態で川に入ってしまったら、袋から出られないカメはおぼれてしまうので、気を付ける。カメを入れた容器を締め切った車の中に置き去りにしてはいけない。直射日光のあたる日向に長時間おきっぱなしにしてもいけない。どちらの場合もカメを熱死させてしまう危険がある。
3.カメの測定、年齢の判定
実験室かベースキャンプに持ち帰ったカメには、種の判別、測定、個体識別のためのマーキングを施す。測定・記録・ウィ[終えたカメは可能な限り早く捕獲地点に戻して放逐する。
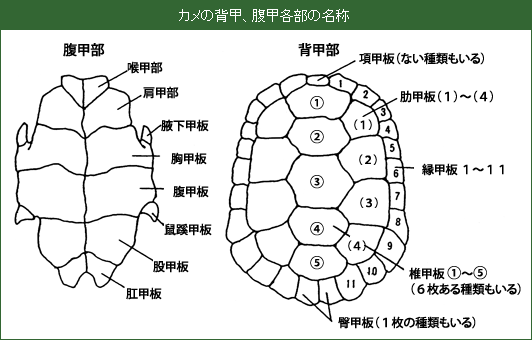
カメの種類を同定することは簡単なようで難しい。もともと日本本土に生息する淡水性のカメは3種類しかいないので、この3種類(イシガメ、クサガメ、スッポン)の見分け方は図鑑で容易に学ぶことができる。難しい点は、イシガメがみつかることが多くなっている点である。クサガメの特徴は背甲(背中の甲羅)に正中線上とその左右に2本、合わせて3本のキールが発達していることだ。イシガメは、背甲後部の縁甲板が鋸の歯のようにギザギザしていることで、クサガメの区別ができる。しかし、地域によって、両方の特徴を併せ持ったカメが捕獲されることがある。その場合は、甲羅以外の特徴を詳しく調べ、体のどの部分がイシガメで、どの部分がクサガメの特徴を示しているのか、詳しく記録する必要がある。
![]() 雌雄の判定: カメの性別判定は、成熟した個体であれば比較的簡単である。腹甲(腹の甲羅)が、雄ではわずかにくぼんでいる。交尾の際に、雌の背に乗りかかるので、雄の腹甲がくぼんでいることによって交尾姿勢を安定させることができる。さらに、総排泄孔(肛門)の位置が成熟した雄と雌では異なる。腹甲の後端から尾の先端の長さを100とすると、雄の総排泄孔の位置は腹甲の後端から60くらいになるが、雌では40程度となる。別の見方では、雄の総排泄孔は背甲の後端よりも後ろに位置するが、雌では背甲の後端よりも内側になる。これは雄の尾の基部にペニスが格納されているためであり、総排泄孔の位置が後ろにあるほど勃起したペニスを雌の総排泄孔に挿入しやすくなることから生じた2次特性ということができる。
雌雄の判定: カメの性別判定は、成熟した個体であれば比較的簡単である。腹甲(腹の甲羅)が、雄ではわずかにくぼんでいる。交尾の際に、雌の背に乗りかかるので、雄の腹甲がくぼんでいることによって交尾姿勢を安定させることができる。さらに、総排泄孔(肛門)の位置が成熟した雄と雌では異なる。腹甲の後端から尾の先端の長さを100とすると、雄の総排泄孔の位置は腹甲の後端から60くらいになるが、雌では40程度となる。別の見方では、雄の総排泄孔は背甲の後端よりも後ろに位置するが、雌では背甲の後端よりも内側になる。これは雄の尾の基部にペニスが格納されているためであり、総排泄孔の位置が後ろにあるほど勃起したペニスを雌の総排泄孔に挿入しやすくなることから生じた2次特性ということができる。
このように、成熟した個体については、雌雄の判定は外形的特徴から容易に行うことができるが、未成熟個体については難しい。解剖して、生殖腺を調べれば確実に雌雄判定できるが、そのようにして判定した標本の外形を見直してみても、雌雄の判定は困難である。
![]() 年齢査定: カメは外形的特徴から年齢査定を行うことの出来る希有な動物である。カメの甲羅に形成される年輪が非活動期に形成される成長停止線とみなすことができるため、この年輪の数を数えることによって、カメが何度成長を止めたかがわかるのである。一般に成長が止まるのは水温が下がってカメが冬眠する冬であるとみなすことができる。つまり、年輪の数はカメが冬を越した回数を示すとされている。
年齢査定: カメは外形的特徴から年齢査定を行うことの出来る希有な動物である。カメの甲羅に形成される年輪が非活動期に形成される成長停止線とみなすことができるため、この年輪の数を数えることによって、カメが何度成長を止めたかがわかるのである。一般に成長が止まるのは水温が下がってカメが冬眠する冬であるとみなすことができる。つまり、年輪の数はカメが冬を越した回数を示すとされている。
温帯のカメは初夏に産卵し、卵は秋に孵化する。そのため、孵化直後の個体には成長の停止を示す年輪はない。この子ガメが越冬し、次の成長期間に十分な成長をすることで年輪が1本形成される。同じようにして2年目に2本、3年目に3本、と増えていく。しかし、冬を越したばかりの個体では、成長停止線の数は実際に越した冬の回数よりも少なくなっている。そのため、年輪の数を数えて単純に年齢とすることはできない。年輪を読む時期、タイミングの情報が必要なのである。そこで、カメの年齢を記録する場合には、X才とだけ記入するよりも(年齢を満年齢か、かぞえ年とするかを区別しないと、後で非常に混乱する)、年輪をさかのぼって生まれた年の絶対年を記入する方が正確な記述となる。
![]() 測定: 体の大きさを、背甲あるいは腹甲の長さを用いて表現する。想定はノギスを用い、0.1mm単位で測定する。体重はよく水を拭き取ったうえで電子天秤やバネばかりを用いて秤量する。
測定: 体の大きさを、背甲あるいは腹甲の長さを用いて表現する。想定はノギスを用い、0.1mm単位で測定する。体重はよく水を拭き取ったうえで電子天秤やバネばかりを用いて秤量する。
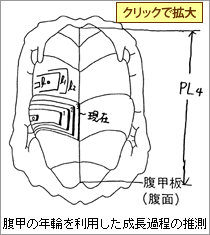
4.腹甲の年輪を利用した成長過程の推測
2004年の6月に捕獲した個体が下図のような年輪を持っていた場合には、2000年が孵化年で、現在4年目と数える。年齢は4才(2000年)と表記するとよい。
 |
 |
5.個体識別
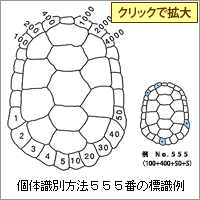 一度捕獲した個体を何らかの方法で標識しておけば、次に捕獲したときに同一個体であることを確認することができる。そうすることで、成長の様子、成熟する年齢、行動範囲など貴重な情報を得ることができる。これを標本再捕獲法による成長、個体群解析と称する。カメの個体識別に用いる標識は、背甲の縁甲板にドリルで穴を開ける方法が一般的で、番号付けは右図の方式を採用している。
一度捕獲した個体を何らかの方法で標識しておけば、次に捕獲したときに同一個体であることを確認することができる。そうすることで、成長の様子、成熟する年齢、行動範囲など貴重な情報を得ることができる。これを標本再捕獲法による成長、個体群解析と称する。カメの個体識別に用いる標識は、背甲の縁甲板にドリルで穴を開ける方法が一般的で、番号付けは右図の方式を採用している。
右図の例では、100番、400番、50番、5番の位置の縁甲板にドリルで穴をあけたので、それぞれの番号を合計して555番がこのカメの個体番号として与えられる。
6.個体数の推定
今回の調査でも、捕獲した全てのカメに標識を施す。御原川のある区域に何匹のカメが生息しているのか、河川の一定域内(約1km)にワナをかけて、カメの生息個体数を推定する。
| 基本推定式(Lincoln-Petersen Index) |  |
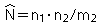 |
|
| 1)サンプル(捕獲個体)数が少ない時でかつ生息個体数に対してサンプル数が小さい時(Baileyの修正式) | |
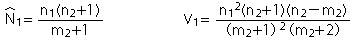 |
|
| 2)サンプル(捕獲個体)数が少ない時でかつ生息個体数に対してサンプル数が大きい時(Chapmanの修正式) | |
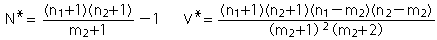 |
![]() データの解析方法: 10日のセミナー後、データシートの作成後、各自相談の上で、グラフ用紙に取りまとめの図、表の形式を試作する。それをもとにレポートの構成、取り入れる図表を絞り込む。最終的なレポートの出来上がりをイメージし、野外実習最終日前日の夜までに、レポート用紙に仮割り付けを作成する。
データの解析方法: 10日のセミナー後、データシートの作成後、各自相談の上で、グラフ用紙に取りまとめの図、表の形式を試作する。それをもとにレポートの構成、取り入れる図表を絞り込む。最終的なレポートの出来上がりをイメージし、野外実習最終日前日の夜までに、レポート用紙に仮割り付けを作成する。
参考
1.カメの生息環境としての河川環境測定項目
- 流速 : ピンポン玉またはスーパーボールを複数流してストップウォッチで時間測定
- 水温、気温
- 川の林冠鬱閉度 : 目測で林冠鬱閉度を%で記録
- 底質 : 川を横断すると卵セクトを複数設置し、岸から岸までの間で底質の長さとその底質の始まり、中間、終わりの水深を記録する
- 底質の分類 : 泥、シルト、砂、れき、岩盤
- 河川構造のタイプ : 瀬、淵、滝など
- カバー構造 : 水辺の植生、倒木、浮石など
- 川岸の植生 : ワナを設置した位置の上流と下流側10mの樹木の種類と樹高を記録
- 川幅 : 水の流れている部分と河原幅、最大増水時の川幅、水深
- 位置 : 千葉県メッシュ番号、市町村名、大字、小字、河川名、最寄の橋名、GPSの緯度経度
2.カメの外部形態測定項目
- 通し番号
- 調査毎の捕獲通し番号
- 調査(捕獲)日
- 捕獲(ワナかけ)場所の地名、河川名
- 捕獲(ワナかけ)場所の軽度・緯度
- 個体(識別)番号
- 種名
- 性別
- 黒化の段階
- 年齢(孵化の年)
- 体重
- 背甲長
- 腹甲長
- 背甲副長
- 背甲副長を計測した縁甲板の番号
- 頭副長
- 尾長
- 体の特徴、甲羅の傷、寄生虫など
- 発見時の行動
- 備考