
定温脊椎動物ではメラノサイトがほとんど唯一の色素細胞であり、ここで生産された色素、メラニンを含む顆粒であるメラノソームが表皮の角化細胞に移行するので、皮膚色の濃さは含まれるメラニン量で決まる。
変温脊椎動物や無脊椎動物の色素細胞も色素物質を生産するが、一般的にはこれらを分泌せずに引き続き含有しており、定温脊椎動物の色素細胞と区別して「色素胞」と呼ばれることが多い。また、色素を含んでいないものの、光をよく反射する結晶を含み、反射された光で色を発現する細胞も色素胞と呼ばれている。
黒い色素メラニンを生産・含有する色素は黒色素胞、カロテノイド(餌から摂取)やプテリジンを含有して赤や黄色を呈色する細胞は赤色素胞・黄色素胞と称され、通常は真皮に存在する。いずれも枝状突起を皮膚の平面に平行に放射状に伸ばした形をとっており、これは発生学的に神経細胞の仲間であることと無縁ではなさそうである。神経細胞の樹状突起は三次元的に伸びるが、黒色素胞の場合でも、枝状部を体表側に曲げ、他の色素胞種を包み込む立体的な構成を取る場合もある(熱帯の海産魚ルリスズメダイや両生類・は虫類の黒色素胞)。これらの色素胞に含まれる色素は光を吸収するので、光吸収性色素胞と総称される。
色素胞の呈色にあずかる色素は、1重(単位膜)の限界膜に囲まれた細胞小器官(色素顆粒と呼ばれる)の基質に含まれている。
さらに色素胞内では、細胞体のほぼ中央部の微小管構成中心から枝状部末端まで、多数の微小管が放射状に伸びており、色素顆粒はこの微小管をレールとして、細胞体中心部へ、あるいは枝状部末端へ向けて移動する。
色素顆粒が細胞中心部に凝集すると、皮膚面で色の占める面積が減少して色調が薄れる。 逆に枝状部を含めた細胞全体に拡散すると、色の面積が増し、色彩が増強される。 この間、細胞の輪郭は変わらない(→図1A)。
色素顆粒の凝集・拡散運動の実態は、神経の軸索輸送同様、微小管の構成蛋白質、チューブリンと、色素顆粒とチューブリンの間に入り込んだモーター蛋白質の細胞質ダイニン(凝集)、あるいはキネシン(拡散)との相互作用である。モーター蛋白質のATPase活性を介して、移動のための運動エネルギーが生じる。
メダカ目の魚類や海産魚メバルの真皮に存在する白色素胞は、実際は色素物質を含んでいないが、広い波長域の光を散乱する細胞小器官を含み、数は少ないものの、枝状突起を有する細胞である。 透過光型顕微鏡では黒色素胞との区別がつきにくいが、落射照明下では白く見えるので白色素胞と称される。 光散乱性の細胞小器官も凝集・拡散するが、その反応は光吸収性色素細胞ほど早くないので、運動メカニズムが全く同じであるかどうかについては更に検討する必要がある。
白色素胞の反応は黒色素胞などとは逆の場合がある。 例えば、交感神経伝達物質のノルエピネフリン(norepinephrine,NE)あるいは血管内皮細胞から分泌されるエンドセリンに対して、光吸収性の色素胞では色素顆粒の凝集が、白色素胞では拡散が生じるが、この現象は皮膚の明暗変化を効率的に進行させる。 黒色素胞と白色素胞がそれぞれNEやエンドセリンに対して異なるタイプの受容体をもつことによって、こうした反応性の違いが生じるのである。
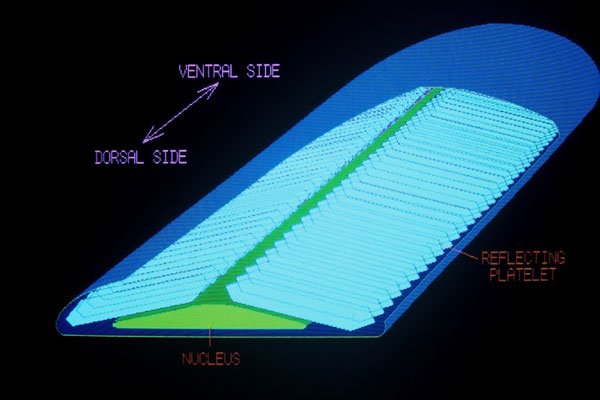
虹色素胞は通常は突起を持たず、楕円状あるいは多角体状の光反射性色素胞である。 色素を含まないが、その細胞内には主としてグアニンからなる板状結晶(光反射小板;無色透明)の重なり(小板堆)が存在する。 屈折率の高いプリン結晶体の重層で生じる反射光の薄膜干渉現象によって、鏡のような高い光反射性(タチウオ、サンマ、カツオなどの体側・腹部の銀白色)や蛍光色のような鮮やかな色合い(ルリスズメダイのコバルトブルー、ネオンテトラ縦縞の青緑色など)が生じる。
前者に係わる虹色素胞内のプリン結晶は厚く、大きく、その規則的成層による光反射性は重層薄膜干渉の“理想型”である。実際、高屈折率(反射小板;無色透明)と低屈折率(細胞質)の光学的厚さ(屈折率×実際の厚さ)が等しくなるように重なった小板堆が、電子顕微鏡によっても観察されている。
重層の構成条件がこれから外れると、干渉は後者の“非理想型”となる。 多くの場合、グアニン結晶の実際の厚みが極めて薄くなり、細胞質と反射小板の光学的厚さが著しく異なるようになる。
 例えば、ルリスズメダイの場合、反射小板の実際の厚みは5nmと薄く、それに対して細胞質の実際の厚みは100nmか、それ以上に達する。
こうした非理想型の場合、反射光スペクトルの主ピークが低下し、ピーク幅が狭くなるという特徴を示す。有色の物質がないのに非常に鮮やかな色彩が発現し、“構造色”、“物理色”などと呼ばれる。
例えば、ルリスズメダイの場合、反射小板の実際の厚みは5nmと薄く、それに対して細胞質の実際の厚みは100nmか、それ以上に達する。
こうした非理想型の場合、反射光スペクトルの主ピークが低下し、ピーク幅が狭くなるという特徴を示す。有色の物質がないのに非常に鮮やかな色彩が発現し、“構造色”、“物理色”などと呼ばれる。
非理想型では、反射率上昇のためには反射小板の枚数を十分に増加させることが必要で、ネオンテオトラ縦縞部の虹色素胞には100枚〜150枚のグアニン結晶が重層した小板堆が2列配列されている。
 理想型の薄膜重層を持つ虹色素胞は一般的に運動性を欠いている。非理想型に属し、青や黄色を呈する虹色素胞では運動性を持つものが多いが、持たないものもある。
スズメダイ科、テトラ類、ニザダイ科のナンヨウハギなどの虹色素胞は運動性を有し、体色変化において重要な役割を演じている。
理想型の薄膜重層を持つ虹色素胞は一般的に運動性を欠いている。非理想型に属し、青や黄色を呈する虹色素胞では運動性を持つものが多いが、持たないものもある。
スズメダイ科、テトラ類、ニザダイ科のナンヨウハギなどの虹色素胞は運動性を有し、体色変化において重要な役割を演じている。
ルリスズメダイやナンヨウハギの運動性虹色素胞内では、細胞の皮膚表面側に存在する核領域から反射小板堆が放射状に配列しており、この堆を構成する小板の間隔が並行的に一斉に変化することが運動の実体で、その結果、反射光のスペクトルが移動する。 間隔の増大・減少はスペクトルのピークをそれぞれ長波長・短波長側に移動させる(→図1B)。 この変化はわずか1〜2分で起こる。小板の間隔が変化する機構は未だに明確になっていないが、チューブリン−ダイニン系が関与されると推察されている。
|
電子顕微鏡でみたナンヨウハギの虹色素胞(左)と
光反射小板(赤矢印)の重なり(小板堆)拡大写真(右) |
 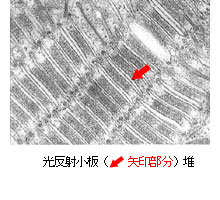 |
デバスズメダイの皮膚にも類似の虹色素胞が見られるが、ここでは黒色素胞の細胞体の上に存在する虹色素胞だけが運動性を持ち、周辺の虹色素胞はいつも黄色を呈していて、運動性を欠いている。運動性虹色素胞が青色を反射しているときは、周辺の黄色を反射する虹色素胞との共存が点描効果を生じ、皮膚は緑色に見える。
 ネオンテトラ体側中央の縦縞部にびっしりならんだ虹色素胞内には2列の光反射小板堆が存在し、小板の間隔は、ヴェネチアン・ブラインド式とも言える機構により増大・減少する(→図1C)。すなわち反射小板の傾きが一斉に変化することにより間隔も変化し、反射光は紫から朱色まで連続的に変わる。
この傾斜角度の変化にはチューブリンとダイニンの相互作用のほか、アクチンの関与の可能性が指摘されている。
ネオンテトラ体側中央の縦縞部にびっしりならんだ虹色素胞内には2列の光反射小板堆が存在し、小板の間隔は、ヴェネチアン・ブラインド式とも言える機構により増大・減少する(→図1C)。すなわち反射小板の傾きが一斉に変化することにより間隔も変化し、反射光は紫から朱色まで連続的に変わる。
この傾斜角度の変化にはチューブリンとダイニンの相互作用のほか、アクチンの関与の可能性が指摘されている。
![]() ドンコ型運動性虹色素胞
ドンコ型運動性虹色素胞
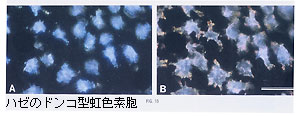 ドンコなどのハゼ科魚類では、枝状部をもち、小型の光反射小板が凝集・拡散するという、白色素胞と似たタイプの虹色素胞が報告された。反射小板が細胞内に広く分布しているときには、小板は比較的ランダムな配向を示し、反射光も白色から黄味がかった色に見える(写真B)。小板が細胞中心部に凝集するにしたがって小板集団が青く見えるようになる(写真A)。
これは、凝集により次第に明瞭な小板堆が形成されていき、重層薄膜干渉現象による反射スペクトル・ピークが成長しつつ、小板間隔の減少に伴って短波長側へ移動、青味掛かった干渉色が現れることによる(→図1D)。両生類の虹色素胞も枝状部を有しており、光反射についても同様の光学機構が当てはまるであろう。
ドンコなどのハゼ科魚類では、枝状部をもち、小型の光反射小板が凝集・拡散するという、白色素胞と似たタイプの虹色素胞が報告された。反射小板が細胞内に広く分布しているときには、小板は比較的ランダムな配向を示し、反射光も白色から黄味がかった色に見える(写真B)。小板が細胞中心部に凝集するにしたがって小板集団が青く見えるようになる(写真A)。
これは、凝集により次第に明瞭な小板堆が形成されていき、重層薄膜干渉現象による反射スペクトル・ピークが成長しつつ、小板間隔の減少に伴って短波長側へ移動、青味掛かった干渉色が現れることによる(→図1D)。両生類の虹色素胞も枝状部を有しており、光反射についても同様の光学機構が当てはまるであろう。
これらの虹色素胞内における反射小板の凝集・拡散は1〜2時間かけて非常にゆっくりと進行するので、反射小板の間隔の変化により1〜2分の内に物理色が変わるタイプの虹色素胞とはその運動メカニズムが異なるものと推察される。 枝状部細胞質表層に存在するアクチンフィラメントの収縮・弛緩により、小板の凝集・拡散が生じている可能性がある。
![]()