 Enami乮1955乯偑偦偺懚嵼傪巜揈丄偦偺屻Baker傜偵傛偭偰惛椡揑偵尋媶偝傟丄1983擭丄Kawauchi傜偵傛偭偰堦師峔憿偑寛掕偝傟偨儊儔僯儞嬅廤儂儖儌儞乮MCH乯偼17屄偺傾儈僲巁偐傜惉傞娐忬偺儁僾僠僪偱偁傞丅峈懱傪梡偄偰偺柶塽慻怐壔妛揑専嶕偐傜丄帇彴壓晹偺恄宱暘斿嵶朎偱嶻惗偝傟丄嫑椶偱偼屻梩偵塣偽傟偰暘斿偝傟傞偙偲偑傢偐偭偨丅
Enami乮1955乯偑偦偺懚嵼傪巜揈丄偦偺屻Baker傜偵傛偭偰惛椡揑偵尋媶偝傟丄1983擭丄Kawauchi傜偵傛偭偰堦師峔憿偑寛掕偝傟偨儊儔僯儞嬅廤儂儖儌儞乮MCH乯偼17屄偺傾儈僲巁偐傜惉傞娐忬偺儁僾僠僪偱偁傞丅峈懱傪梡偄偰偺柶塽慻怐壔妛揑専嶕偐傜丄帇彴壓晹偺恄宱暘斿嵶朎偱嶻惗偝傟丄嫑椶偱偼屻梩偵塣偽傟偰暘斿偝傟傞偙偲偑傢偐偭偨丅
曄壏愐捙摦暔偺旂晢偺怓慺朎偼丄儂儖儌儞偵傛偭偰傕暋嶨偵僐儞僩儘乕儖偝傟偰偄傞丅 嵟嬤偺抦尒偼塿乆偦偺暋嶨偝傪報徾偯偗傞乮仺恾俀乯丅擼壓悅懱拞梩儂儖儌儞乮兛-MSH乯偼曄壏愐捙摦暔偺怓慺朎慡斒偵岠壥偑偁傝丄堦斒偵怓慺梓棻傗敀怓慺朎撪偺岝嶶棎惈嵶朎彫婍姱傪嶶棎偝偣傞丅MSH偺嶌梡偵偼丄嵶朎奜偵僇儖僔僂儉僀僆儞偑懚嵼偟偰偄傞偙偲偑昁梫偱偁傞丅僪儞僐偺塣摦惈擑怓慺朎偱偼岝斀幩彫斅偺嬅廤偑婲偙傞丅偄偢傟傕MSH庴梕懱傪夘偟偰嵶朎撪cAMP擹搙偑忋徃偟偨寢壥偱偁傞丅孞傝曉偟弎傋傞偙偲偵側傞偑丄僴僛壢嫑椶偺塣摦惈擑怓慺朎偵偍偗傞彫斅偺嬅廤丒奼嶶偺偟偔傒偼丄懠偺怓慺朎偺婡峔偲堎側傞傜偟偔丄嵶朎撪僙僇儞僪儊僢僙儞僕儍乕偺擹搙偲塣摦偺曽岦偲偺娭學偼憡屳偵斀懳偱偁傞丅
堦曽丄儖儕僗僘儊僟僀宆偺丄偄傢備傞旕棟憐宆廳憌敄枌姳徛尰徾偵傛傞暔棟怓傪惗偠傞擑怓慺朎偼傎偲傫偳儂儖儌儞偺惂屼傪庴偗偢丄傕偭傁傜恄宱巟攝偺傒偑嶌摦偟偰偄傞丅偙傟傜偺暔棟怓偑丄孮傟傪惉偡屄懱娫偺僐儈儏僯働乕僔儑儞偵棙梡偝傟偰偄傞応崌偑懡偔丄憗偄懱怓曄壔偑媮傔傜傟偰偄傞偨傔偐傕偟傟側偄丅
僯僕儅僗傗僂僫僊傪崟偄攚抧偵揔墳偝偣傞偲MSH偺寣拞擹搙偼忋徃偡傞偑丄僸儔儊偺拠娫偱偼曄壔偑側偐偭偨偲偺曬崘偑偁傞丅幚嵺偺攚抧揔墳偵偍偄偰偼丄儂儖儌儞埶懚宆乮慜幰乯偲恄宱埶懚宆乮屻幰乯偑偁傞偺偐傕偟傟側偄丅 MSH偼惗棟妛揑懱怓曄壔偵娭傢傞偩偗偱側偔丄挿婜娫丄攚抧揔墳偝偣傜傟偨応崌偺宍懺妛揑懱怓曄壔偵傕娭梌偡傞丅儌僓儞價乕僋僥傿儔僺傾偵10擔娫埲忋偵傢偨偭偰兛-MSH傪搳梌偟懕偗偨偲偙傠丄攚抧偺怓偵娭學側偔崟怓慺朎偺悢偑憹偊丄僒僀僘傕戝偒偔側偭偨偲偺幚尡寢壥偑偁傞丅
堦掕偺攚抧怓偵惗棟妛揑偵弴墳偟偨忬懺偑偁傞擔悢懕偔偲丄宍懺妛揑側曄壔傕婲偙傞壜擻惈偑峫偊傜傟傞丅MSH偑庴梕懱偵寢崌偡傞偲丄偦偺忣曬偼怓慺梓棻塣摦偺儊僇僲儈僇儖宯偵傕偨傜偝傟傞偙偲偼栜榑偩偑丄摨帪偵妀撪偵傕揱偊傜傟丄堚揱巕偺揮幨挷愡側偳偵傕塭嬁傪梌偊傞偺偱偁傠偆丅
Enami乮1955乯偑偦偺懚嵼傪巜揈丄偦偺屻Baker傜偵傛偭偰惛椡揑偵尋媶偝傟丄1983擭丄Kawauchi傜偵傛偭偰堦師峔憿偑寛掕偝傟偨儊儔僯儞嬅廤儂儖儌儞乮MCH乯偼17屄偺傾儈僲巁偐傜惉傞娐忬偺儁僾僠僪偱偁傞丅峈懱傪梡偄偰偺柶塽慻怐壔妛揑専嶕偐傜丄帇彴壓晹偺恄宱暘斿嵶朎偱嶻惗偝傟丄嫑椶偱偼屻梩偵塣偽傟偰暘斿偝傟傞偙偲偑傢偐偭偨丅
掅擹搙偱偼丄偦偺柤偑帵偡擛偔丄儊儔僯儞梓棻乮崟怓慺梓棻乯偺嬅廤傪桿婲偡傞丅嫑傪敀攚抧偵抲偔偲MCH偺屻梩偐傜偺暘斿傕懀恑偝傟傞丅偟偐偟丄in vitro偺幚尡偱偼丄崅擹搙MCH偼儊儔僯儞梓棻偺奼嶶傪桿婲偡傞応崌偑偁傞丅
椺偊偽丄Synbranchuns marmoratus偲偄偆傾儅僝儞愳偺僂僫僊偺堦庬丄偁傞偄偼僫儅僘壢偺拠娫偺僐儕僪儔僗丒僷儗傾僩僁僗偺旂晢偺崟怓慺朎偼丄MCH偺擹搙偑掅偄偲嬅廤斀墳偡傞偑丄崅擹搙偵側傞偲丄堦扷嬅廤屻丄嵞奼嶶偟丄嵟廔揑側嬅廤棪偼掅偔側傞丅
儊僟僇偺敀怓慺朎傗丄僇僄儖偺崟怓慺朎偱偼丄嬅廤偼尒傜傟偢丄奼嶶偺傒偑惗偠傞丅偙偺奼嶶偵偼奜塼拞偺僇儖僔僂儉僀僆儞偑昁梫偱偁傞丅愒怓慺朎傗墿怓慺朎偺応崌偼丄掅擹搙偱嬅廤丒崅擹搙偱奼嶶偡傞僞僀僾丄柍斀墳僞僀僾丄奼嶶偩偗偡傞僞僀僾側偳丄斀墳惈偼懡條偱偁傞丅MCH庴梕懱偵偼偍偦傜偔擇偮偺僒僽僞僀僾偑偁傝丄戞侾偺僞僀僾偼嫑椶怓慺朎偵堦斒偵懚嵼偟丄掅擹搙偱偺嬅廤斀墳傪拠夘偡傞丅戞俀偺僞僀僾偼斾妑揑崅擹搙偺MCH偵傛偭偰巋寖偝傟傞庴梕懱偱丄僇僄儖崟怓慺朎丄儊僟僇敀怓慺朎丄堦晹偺嫑椶崟怓慺朎丄愒怓慺朎丄墿怓慺朎側偳偵暘晍偟偰偄傞偲悇嶡偝傟傞丅
嵟嬤MCH偼歁擕摦暔偱偼怘梸拞悤傪巋寖偡傞偙偲偑傢偐傝丄懱怓曄壔偲偼柍娭學側偲偙傠偱恖乆偺擬偄帇慄傪梺傃傞傛偆偵側偭偨丅
嫑椶偺摢捀晹丄偪傚偆偳嵍塃偺懁娽偺恀拞偁偨傝偵怓慺朎偺寚擛偟偨晹埵偑偁傝丄偦偺晹暘偺旂晢偺壓偵徏壥懱偲偄偆撪暘斿婍姱偑偁傞丅徏壥懱偺姶妎嵶朎偼栐枌偺帇嵶朎偵椶帡偺峔憿傪桳偟丄奜奅偺岝偺捈愙塭嬁壓偵偁傞丅 偙偙偱崌惉偝傟傞儂儖儌儞偑儊儔僩僯儞偱丄崌惉丒暘斿偼栭偵峴傢傟傞丅 廬偭偰丄拫栭偺儕僘儉偵峀偔娭梌偟偰偄傞壜擻惈偑偁傞丅
儊儔僩僯儞偲偄偆柤偼丄僂僔偺徏壥懱偐傜拪弌偟偨暔幙偑僇僄儖偺旂晢傪柧壔偝偣偨乮Lerner傜丄1958乯偙偲偵桼棃偡傞偑丄嫑椶偺岝媧廂惈怓慺朎偱傕捠忢丄怓慺梓棻偺嬅廤傪桿婲偡傞丅偟偐偟丄嫑庬偵傛偭偰丄偁傞偄偼摨堦懱偱傕晹埵偵傛偭偰儊儔僩僯儞偺怓慺朎偵懳偡傞岠壥偵堘偄偑偁傞偙偲偑傢偐傝丄偙偺儂儖儌儞偲柾條宍惉偲偺娭楢偑巜揈偝傟傞傛偆偵側偭偨丅
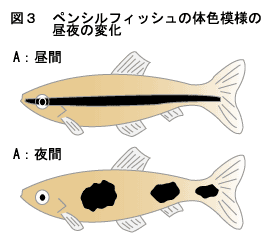 椺偊偽丄儁儞僔儖僼傿僢僔儏偺懱懁偺廲幦偼丄栭偵側傞偲撈摿偺僗億僢僩柾條偵曄傢傞丅栭娫偵僗億僢僩偲偟偰巆偭偰偄傞強偺崟怓慺朎偼丄儊儔僩僯儞偵斀墳偟側偄僞僀僾偱偁傞丅偟偐偟丄傛傝徻嵶偵娤嶡偡傞偲丄栭娫偵惗偠傞僗億僢僩偺捈宎偑拫娫偺廲幦偺暆偵斾傋偰堦抜偲戝偒偄偙偲偵婥晅偔乮恾3乯丅偮傑傝丄栭娫偺僗億僢僩柾條偵娭梌偟偰偄傞崟怓慺朎偺拞偵偼丄拫娫偼嬅廤偟偰偄傞乮棟桼偼晄柧乯偺偵丄栭偵側傞偲儊儔僩僯儞偱奼嶶偡傞傕偺偑偁傝丄偦傟傜偺崟怓慺朎偑怴偨偵奼嶶偟偨寢壥丄戝偒側僗億僢僩偵側偭偨偲峫偊傜傟傞丅
椺偊偽丄儁儞僔儖僼傿僢僔儏偺懱懁偺廲幦偼丄栭偵側傞偲撈摿偺僗億僢僩柾條偵曄傢傞丅栭娫偵僗億僢僩偲偟偰巆偭偰偄傞強偺崟怓慺朎偼丄儊儔僩僯儞偵斀墳偟側偄僞僀僾偱偁傞丅偟偐偟丄傛傝徻嵶偵娤嶡偡傞偲丄栭娫偵惗偠傞僗億僢僩偺捈宎偑拫娫偺廲幦偺暆偵斾傋偰堦抜偲戝偒偄偙偲偵婥晅偔乮恾3乯丅偮傑傝丄栭娫偺僗億僢僩柾條偵娭梌偟偰偄傞崟怓慺朎偺拞偵偼丄拫娫偼嬅廤偟偰偄傞乮棟桼偼晄柧乯偺偵丄栭偵側傞偲儊儔僩僯儞偱奼嶶偡傞傕偺偑偁傝丄偦傟傜偺崟怓慺朎偑怴偨偵奼嶶偟偨寢壥丄戝偒側僗億僢僩偵側偭偨偲峫偊傜傟傞丅
偡側傢偪丄儁儞僔儖僼傿僢僔儏偺廲幦晹偵偼儊儔僩僯儞偵懳偟偰嬅廤偡傞崟怓慺朎丄奼嶶偡傞崟怓慺朎偑嫟懚偟偰偄傞偙偲偵側傞丅
側偍丄儁儞僔儖僼傿僢僔儏偺栭娫偺僗億僢僩偼丄愳掙偺彫愇偵帡偨柾條偵偡傞偙偲偱丄栭峴惈偺曔怘幰偵懳偡傞僇儉僼儔乕僕儏偺岠壥偑偁傞偺偱偼側偐傠偆偐丅
傑偨丄僱僆儞僥僩儔傗僇乕僕僫儖僥僩儔偺愒偄暊晹偵懚嵼偡傞愒怓慺朎傕儊儔僩僯儞偵傛偔斀墳偟丄栭偵側傞偲怓慺梓棻偺嬅廤偵傛偭偰丄愒偄怓偼栚棫偨側偔側傞丅廲幦晹偺擑怓慺朎偼儊儔僩僯儞偺庴梕懱傪帩偨側偄偑丄奜奅偺岝偵捈愙斀墳偡傞僞僀僾側偺偱丄埫強偱偼丄擹偄巼怓偵曄傢傞丅拫娫偺慛傗偐側懱怓偲偼偆偭偰曄傢偭偰栭娫偼栚棫偨側偄怓偵側傝丄栭峴惈曗怓幰偺栚傪摝傟偰偄傞丅
偙偺傛偆偵丄摦暔偼惗偒傞偨傔偵怓傗柾條傪岻柇偵曄壔偝偣傞丅恄宱揱払暔幙偺傒側傜偢丄條乆側儂儖儌儞偺庴梕懱傪偆傑偔慻傒崌傢偣偰丄嬌傔偰暋嶨側怓偺惂屼傪壜擻偵偟偰偄傞偺偱偁傞丅
暃恡悜幙儂儖儌儞偱偁傞僄僺僱僼儕儞偼兝宆傾僪儗僫儕儞庴梕懱偵崅偄恊榓惈傪帩偪丄怓慺梓棻偺奼嶶傪桿婲偡傞丅傑偨丄擼壓悅懱慜梩偐傜暘斿偝傟傞僾儘儔僋僠儞偼丄嫑椶偺愒怓慺朎偲墿怓慺朎偵嶌梡偟偰丄怓慺奼嶶傪堷偒婲偙偡丅 僫僀儖僥傿儔僺傾傗僞僀儕僋僶儔僞僫僑側偳偱偼丄斏怋婜偺梇偵愒偄崶堶怓偑尰傟傞偑丄偦偺愒偄晹埵偵弌尰偡傞愒怓慺朎傗墿怓慺朎偺僾儘儔僋僠儞偵懳偡傞斀墳惈偼嬌傔偰崅偄丅
MSH偵傛偭偰傕怓慺奼嶶偼婲偙傞偑丄偦偺応崌丄崟怓慺朎傕奼嶶偡傞偺偱懱怓偼傓偟傠崟偢傓丅偦偺揰丄僾儘儔僋僠儞偱偁傟偽丄愒丒墿怓慺朎偺傒偱怓慺梓棻偺奼嶶偑桿婲偝傟傞偺偱丄愒怓傪栚棫偨偣傞偵偼搒崌偑傛偄丅
僾儘儔僋僠儞埲奜偺儂儖儌儞庴梕懱偼枌7夞娧捠宆偱丄偦傟偧傟G抈敀幙乮Gs偁傞偄偼Gi乯偲僇僢僾儕儞僌偟丄cAMP傪擇師儊僢僙儞僕儍乕偵偟偰偄傞丅僾儘儔僋僠儞庴梕懱偼傎擕椶傗捁椶偱偼僠儘僔儞僉僫乕僛寢崌宆庴梕懱偲偟偰擣幆偝傟偰偄傞偑丄嫑椶怓慺朎偱偺忣曬揱払宱楬偵偮偄偰偺徻嵶偼崱屻専摙偝傟傞丅
![]()
丂